- Explicar o papel dos músculos na locomoção
- Tipos de tecido muscular e fibras
- Modelo de contração de filamentos deslizantes
- ATP e Contração Muscular
Explicar o papel dos músculos na locomoção
A contração muscular ocorre quando os sarcómeros se encurtam, à medida que os filamentos grossos e finos deslizam uns sobre os outros, o que se designa por modelo de filamentos deslizantes da contração muscular. O ATP fornece a energia para a formação das pontes cruzadas e para o deslizamento dos filamentos. As proteínas reguladoras, como a troponina e a tropomiosina, controlam a formação das pontes cruzadas. O acoplamento excitação-contração transduz o sinal elétrico doO número de fibras musculares que se contraem determina a força que o músculo inteiro produz.
Objectivos de aprendizagem
- Classificar os diferentes tipos de tecidos e fibras musculares
- Descrever os processos de contração muscular
- Discutir porque é que o ATP é necessário para o movimento muscular
- Identificar o papel do cérebro no movimento muscular
- Discutir a tensão e a contração muscular
Tipos de tecido muscular e fibras
As células musculares são especializadas para a contração. Os músculos permitem movimentos como andar e também facilitam processos corporais como a respiração e a digestão. O corpo contém três tipos de tecido muscular: músculo esquelético, músculo cardíaco e músculo liso (Figura 1).
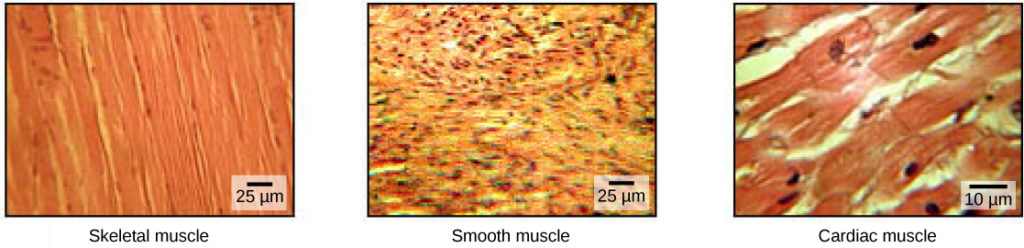
Figura 1. O corpo contém três tipos de tecido muscular: músculo esquelético, músculo liso e músculo cardíaco, visualizados aqui usando microscopia de luz. As células do músculo liso são curtas, afiladas em cada extremidade e têm apenas um núcleo rechonchudo em cada uma. As células do músculo cardíaco são ramificadas e estriadas, mas curtas. O citoplasma pode se ramificar e elas têm um núcleo no centro da célula. (crédito:modificação do trabalho do NCI, NIH; dados da barra de escala de Matt Russell)
Tecido muscular esquelético O músculo esquelético forma os músculos esqueléticos, que se ligam aos ossos ou à pele e controlam a locomoção e qualquer movimento que possa ser conscientemente controlado. Como pode ser controlado pelo pensamento, o músculo esquelético também é chamado de músculo voluntário. Os músculos esqueléticos são longos e de aparência cilíndrica; quando vistos ao microscópio, o tecido muscular esquelético tem uma aparência estriada. As estrias são causadas pora disposição regular das proteínas contrácteis (actina e miosina). Actina é uma proteína contrátil globular que interage com miosina O músculo esquelético também tem vários núcleos presentes numa única célula.
Tecido muscular liso O músculo liso não tem estrias, não está sob controlo voluntário, tem apenas um núcleo por célula, é afilado em ambas as extremidades e é designado por músculo involuntário.
Tecido muscular cardíaco Tal como o músculo esquelético, o músculo cardíaco é estriado, mas ao contrário do músculo esquelético, o músculo cardíaco não pode ser conscientemente controlado e é chamado músculo involuntário. Tem um núcleo por célula, é ramificado e distingue-se pela presença de discos intercalares.
Estrutura das fibras do músculo esquelético
Cada fibra muscular esquelética é uma célula muscular esquelética. Estas células são incrivelmente grandes, com diâmetros de até 100 µm e comprimentos de até 30 cm. A membrana plasmática de uma fibra muscular esquelética é chamada de sarcolema O sarcolema é o local de condução do potencial de ação, que desencadeia a contração muscular. Dentro de cada fibra muscular existem miofibrilhas As miofibrilas percorrem todo o comprimento da fibra muscular e, como têm apenas cerca de 1,2 µm de diâmetro, podem encontrar-se centenas a milhares no interior de uma fibra muscular. Ligam-se ao sarcolema nas suas extremidades, pelo que, quando as miofibrilas encurtam, toda a célula muscular se contrai (Figura 2).
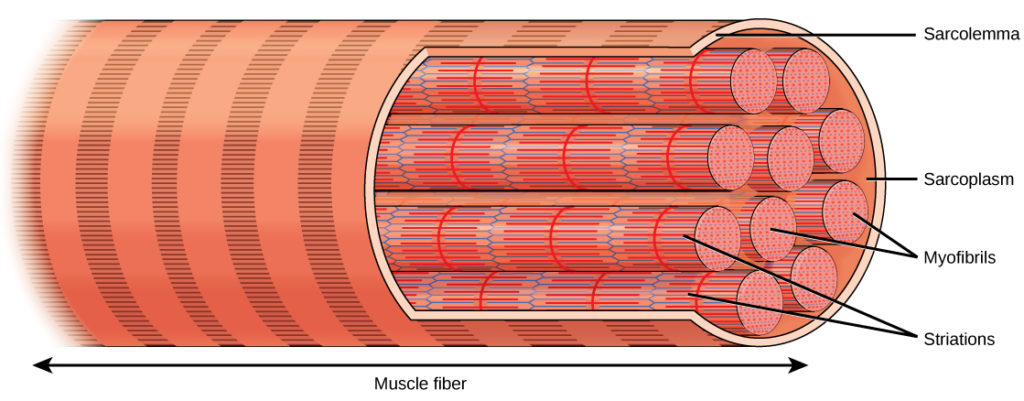
Figura 2. Uma célula muscular esquelética está rodeada por uma membrana plasmática chamada sarcolema e por um citoplasma chamado sarcoplasma. Uma fibra muscular é composta por muitas fibrilas, agrupadas em unidades ordenadas.
A aparência estriada do tecido muscular esquelético resulta da repetição de bandas das proteínas actina e miosina que estão presentes ao longo do comprimento das miofibrilas. As bandas A escuras e as bandas I claras repetem-se ao longo das miofibrilas e o alinhamento das miofibrilas na célula faz com que toda a célula pareça estriada ou com bandas.
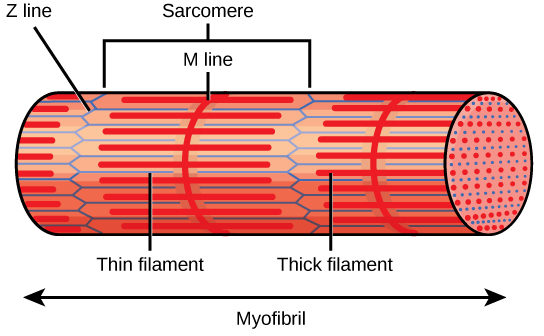
Figura 3. Um sarcómero é a região que vai de uma linha Z até à linha Z seguinte. Estão presentes muitos sarcómeros numa miofibrila, o que resulta no padrão de estriação caraterístico do músculo esquelético.
Cada banda I tem uma linha densa que passa verticalmente pelo meio, denominada disco Z ou linha Z. Os discos Z marcam a fronteira das unidades denominadas sarcómeros Um sarcómero é o espaço entre dois discos Z consecutivos e contém uma banda A inteira e duas metades de uma banda I, uma de cada lado da banda A. Uma miofibrila é composta por muitos sarcómeros ao longo do seu comprimento e, à medida que os sarcómeros se contraem individualmente, as miofibrilas e as células musculares encurtam (Figura 3).
As miofibrilhas são compostas por estruturas mais pequenas chamadas miofilamentos Existem dois tipos principais de filamentos: filamentos grossos e filamentos finos; cada um tem composições e localizações diferentes. Filamentos espessos ocorrem apenas na banda A de uma miofibrila. Filamentos finos ligam-se a uma proteína no disco Z chamada alfa-actinina e ocorrem ao longo de todo o comprimento da banda I e parcialmente na banda A. A região em que os filamentos grossos e finos se sobrepõem tem uma aparência densa, pois há pouco espaço entre os filamentos. Os filamentos finos não se estendem até as bandas A, deixando uma região central da banda A que contém apenas filamentos grossos.A região central da banda A parece ligeiramente mais clara do que o resto da banda A e é chamada de zona H (Figura 4). O meio da zona H tem uma linha vertical chamada linha M, na qual as proteínas acessórias mantêm os filamentos espessos unidos. Tanto o disco Z quanto a linha M mantêm os miofilamentos no lugar para manter o arranjo estrutural e as camadas da miofibrila. As miofibrilas estão conectadas umas às outrasO principal componente dos filamentos finos é a proteína actina. Dois outros componentes dos filamentos finos são a tropomiosina e a troponina. A actina tem sítios de ligação para a ligação da miosina. Os filamentos de tropomiosina bloqueiam os sítios de ligação e impedem as interacções actina-miosina quando os músculos estão em repouso. A troponina é constituída por trêsUma subunidade liga-se à tropomiosina, uma subunidade liga-se à actina e uma subunidade liga-se a iões Ca2+.
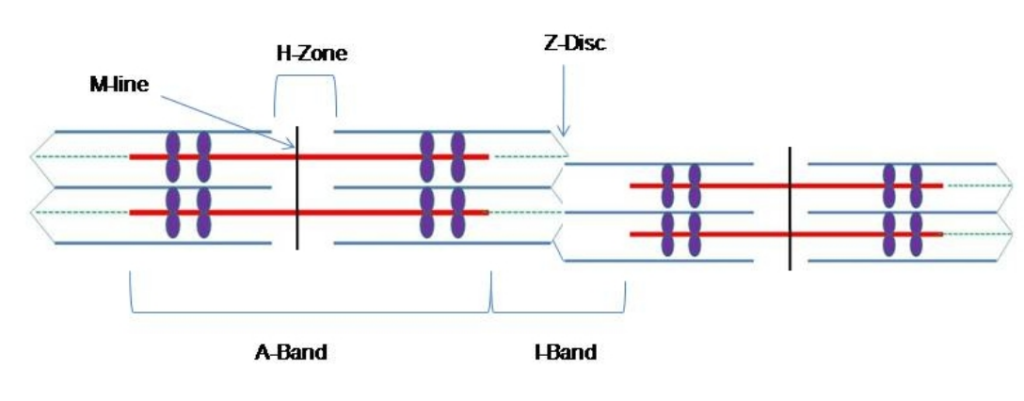
Figura 4 - O sarcómero
Veja este vídeo que mostra a organização das fibras musculares.
Modelo de contração de filamentos deslizantes
Quando um músculo se contrai, a actina é puxada pela miosina em direção ao centro do sarcómero até que os filamentos de actina e miosina se sobreponham completamente. Por outras palavras, para que uma célula muscular se contraia, o sarcómero tem de encurtar. No entanto, os filamentos grossos e finos - os componentes dos sarcómeros - não encurtam. Em vez disso, deslizam uns pelos outros, fazendo com que o sarcómero encurte enquanto os filamentosA teoria dos filamentos deslizantes da contração muscular foi desenvolvida para se ajustar às diferenças observadas nas bandas designadas no sarcómero em diferentes graus de contração e relaxamento muscular. O mecanismo de contração é a ligação da miosina à actina, formando pontes cruzadas que geram o movimento dos filamentos (Figura 5).
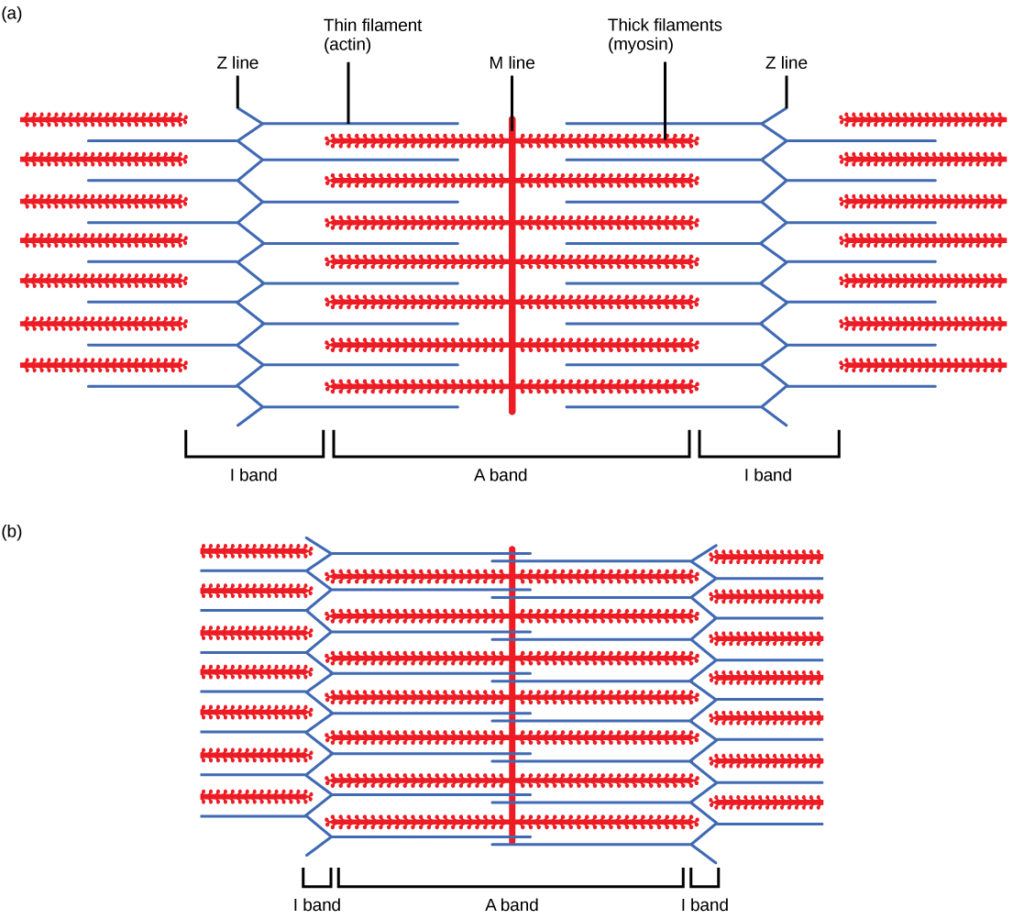
Figura 5. Quando (a) um sarcómero (b) se contrai, as linhas Z aproximam-se e a banda I torna-se mais pequena. A banda A mantém a mesma largura e, na contração total, os filamentos finos sobrepõem-se.
Quando um sarcómero encurta, algumas regiões encurtam enquanto outras mantêm o mesmo comprimento. Um sarcómero é definido como a distância entre dois discos Z consecutivos ou linhas Z; quando um músculo se contrai, a distância entre os discos Z é reduzida. A zona H - a região central da zona A - contém apenas filamentos espessos (miosina) e é encurtada durante a contração. A zona H torna-se cada vez mais pequena devidoAssim, quando o músculo está totalmente contraído, a zona H já não é visível. A banda I contém apenas filamentos finos e também encurta. A banda A não encurta - mantém o mesmo comprimento - mas as bandas A de diferentes sarcómeros aproximam-se durante a contração, acabando por desaparecer. Os filamentos finos são puxadosA zona de sobreposição, na qual os filamentos finos e os filamentos grossos ocupam a mesma área, aumenta à medida que os filamentos finos se deslocam para o interior.
Note-se que os filamentos de actina e miosina não mudam de comprimento, mas deslizam uns sobre os outros.
ATP e Contração Muscular
O movimento de encurtamento muscular ocorre quando as cabeças de miosina se ligam à actina e puxam a actina para dentro. Esta ação requer energia, que é fornecida pelo ATP. A miosina liga-se à actina num local de ligação na proteína globular actina. A miosina tem outro local de ligação para o ATP, no qual a atividade enzimática hidrolisa o ATP em ADP, libertando uma molécula de fosfato inorgânico e energia.
A ligação do ATP faz com que a miosina liberte a actina, permitindo que a actina e a miosina se separem uma da outra. Depois de isto acontecer, o ATP recém-ligado é convertido em ADP e fosfato inorgânico, P i A energia libertada durante a hidrólise do ATP altera o ângulo da cabeça da miosina para uma posição "inclinada". A cabeça da miosina está então em posição de continuar o movimento, possuindo energia potencial, mas o ADP e o P i Se os locais de ligação da actina estiverem cobertos e indisponíveis, a miosina permanecerá na configuração de alta energia com o ATP hidrolisado, mas ainda ligada.
Se os locais de ligação da actina estiverem descobertos, forma-se uma ponte cruzada, ou seja, a cabeça da miosina percorre a distância entre as moléculas de actina e de miosina. P i A cabeça da miosina move-se em direção à linha M, puxando a actina consigo. À medida que a actina é puxada, os filamentos movem-se cerca de 10 nm em direção à linha M. Este movimento é designado por curso de força, uma vez que é o passo em que a força é produzida. À medida que a actina é puxada em direção à linha M, o sarcómero encurtae o músculo contrai-se.
Quando a cabeça da miosina está "inclinada", contém energia e está numa configuração de alta energia. Esta energia é gasta à medida que a cabeça da miosina se move através do curso de força; no final do curso de força, a cabeça da miosina está numa posição de baixa energia. Após o curso de força, o ADP é libertado; no entanto, a ponte cruzada formada ainda está no lugar, e a actina e a miosina estão ligadas entre si. O ATP pode então ligar-se aO movimento da cabeça da miosina de volta à sua posição original é chamado de curso de recuperação. Os músculos em repouso armazenam energia do ATP nas cabeças da miosina enquanto esperam por outra contração.
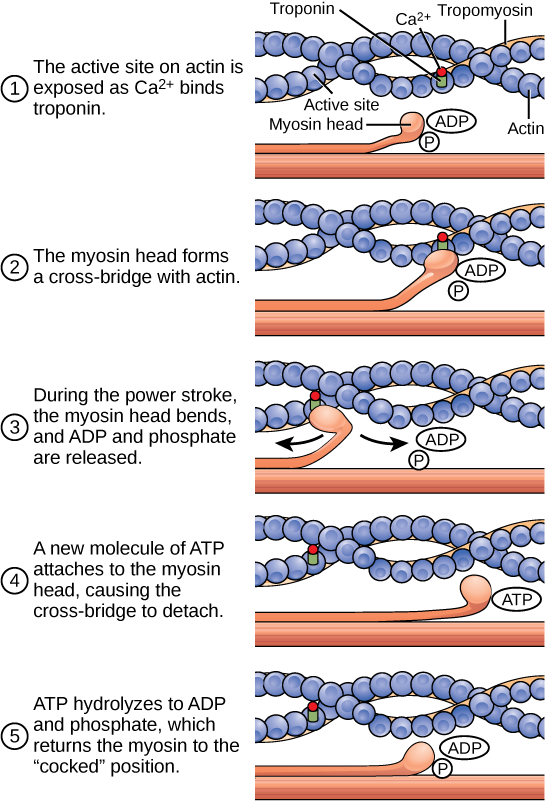
Figura 6: O ciclo de contração muscular de ponte cruzada, que é desencadeado pela ligação do Ca2+ ao local ativo da actina, é mostrado. Com cada ciclo de contração, a actina move-se em relação à miosina.
Proteínas reguladoras
Quando um músculo está em estado de repouso, a actina e a miosina estão separadas. Para impedir que a actina se ligue ao local ativo da miosina, as proteínas reguladoras bloqueiam os locais de ligação molecular. Tropomiosina bloqueia os locais de ligação da miosina nas moléculas de actina, impedindo a formação de pontes cruzadas e a contração de um músculo sem estímulo nervoso. Troponina liga-se à tropomiosina e ajuda a posicioná-la na molécula de actina; também liga iões de cálcio.
Para permitir uma contração muscular, a tropomiosina tem de mudar de conformação, descobrindo o local de ligação da miosina numa molécula de actina e permitindo a formação de pontes cruzadas. Isto só pode acontecer na presença de cálcio, que é mantido em concentrações extremamente baixas no sarcoplasma. Se estiver presente, os iões de cálcio ligam-se à troponina, causando alterações conformacionais na troponina que permitem que a tropomiosina se afasteAssim que a tropomiosina é removida, pode formar-se uma ponte cruzada entre a actina e a miosina, desencadeando a contração. O ciclo da ponte cruzada continua até que os iões Ca2+ e o ATP deixem de estar disponíveis e a tropomiosina volte a cobrir os locais de ligação da actina.
Veja este vídeo que explica como é sinalizada uma contração muscular.