- Identificar os reagentes e produtos da respiração celular e onde estas reacções ocorrem numa célula
- Glicólise
- Oxidação do piruvato
- Ciclo do ácido cítrico
- Cadeia de transporte de electrões
Identificar os reagentes e produtos da respiração celular e onde estas reacções ocorrem numa célula
Agora que aprendemos como os autótrofos, como as plantas, convertem a luz solar em açúcares, vamos ver como todos os eucariotas - o que inclui os humanos - utilizam esses açúcares.
No processo de fotossíntese, as plantas e outros produtores fotossintéticos criam glucose, que armazena energia nas suas ligações químicas. Depois, tanto as plantas como os consumidores, como os animais, passam por uma série de vias metabólicas - coletivamente designadas por respiração celular. A respiração celular extrai a energia das ligações da glucose e converte-a numa forma que todos os seres vivos podem utilizar.
Objectivos de aprendizagem
- Descrever o processo de glicólise e identificar os seus reagentes e produtos
- Descrever o processo de oxidação do piruvato e identificar os seus reagentes e produtos
- Descrever o processo do ciclo do ácido cítrico (ciclo de Krebs) e identificar os seus reagentes e produtos
- Descrever a cadeia respiratória (cadeia de transporte de electrões) e o seu papel na respiração celular
A respiração celular é um processo que todos os seres vivos utilizam para converter glicose em energia. Os autótrofos (como as plantas) produzem glicose durante a fotossíntese. Os heterótrofos (como os seres humanos) ingerem outros seres vivos para obter glicose. Embora o processo possa parecer complexo, esta página apresenta os elementos-chave de cada parte da respiração celular.
Glicólise
Glicólise A glicólise é o primeiro passo na decomposição da glicose para extrair energia para o metabolismo celular. Quase todos os organismos vivos realizam a glicólise como parte do seu metabolismo. O processo não utiliza oxigénio e, por isso, é anaeróbio (A glicólise ocorre no citoplasma das células procarióticas e eucarióticas. A glicose entra nas células heterotróficas de duas formas.
- Através de transporte ativo secundário, em que o transporte se realiza contra o gradiente de concentração de glucose.
- Através de um grupo de proteínas integrais chamadas proteínas GLUT, também conhecidas como proteínas transportadoras de glicose, estes transportadores ajudam na difusão facilitada da glicose.
A glicólise começa com a estrutura em forma de anel de seis carbonos de uma única molécula de glucose e termina com duas moléculas de um açúcar de três carbonos chamado piruvato (Figura 1).
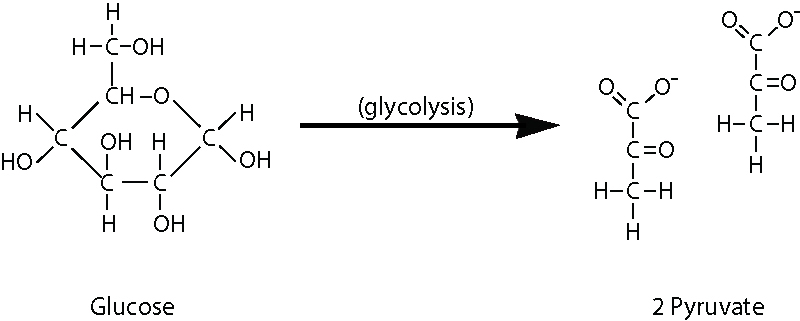
Figura 1: Reactores e produtos da glicólise.
A glicólise é composta por dez etapas divididas em duas metades distintas. A primeira metade da glicólise é também conhecida como etapas que requerem energia Esta via retém a molécula de glicose na célula e utiliza energia para a modificar de modo a que a molécula de açúcar de seis carbonos possa ser dividida uniformemente em duas moléculas de três carbonos. A segunda parte da glicólise (também conhecida como etapas de libertação de energia ) extrai energia das moléculas e armazena-a sob a forma de ATP e NADH, a forma reduzida do NAD.
Primeira metade da glicólise (etapas que requerem energia)
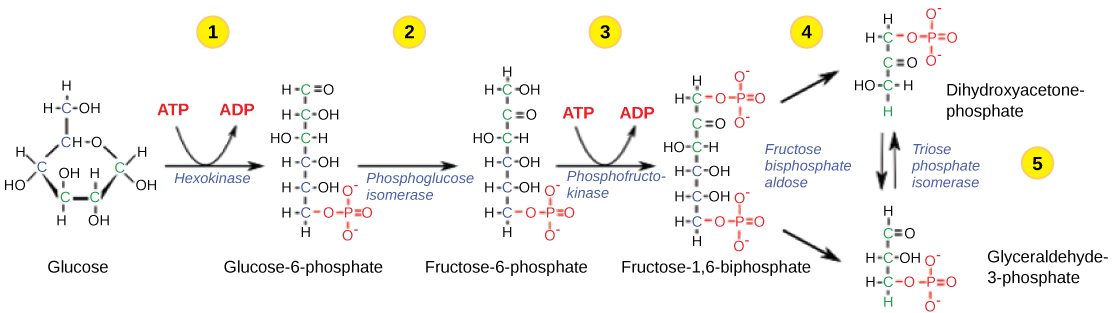
Figura 2. A primeira metade da glicólise utiliza duas moléculas de ATP na fosforilação da glucose, que é depois dividida em duas moléculas de três carbonos.
Etapa 1: A primeira etapa da glicólise é catalisada pela hexoquinase, uma enzima com grande especificidade que catalisa a fosforilação de açúcares com seis carbonos. A hexoquinase fosforila a glicose utilizando ATP como fonte de fosfato, produzindo glicose-6-fosfato, uma forma mais reactiva de glicose. Esta reação impede que a molécula de glicose fosforilada continue a interagir com os GLUTproteínas, e já não pode sair da célula porque o fosfato carregado negativamente não lhe permite atravessar o interior hidrofóbico da membrana plasmática.
Etapa 2: Na segunda etapa da glicólise, uma isomerase converte a glicose-6-fosfato num dos seus isómeros, a frutose-6-fosfato. isomerase A fosfoglucose é uma enzima que catalisa a conversão de uma molécula num dos seus isómeros. Esta mudança de fosfoglucose para fosfofrutose permite a eventual divisão do açúcar em duas moléculas de três carbonos.
Etapa 3: A terceira etapa é a fosforilação da frutose-6-fosfato, catalisada pela enzima fosfofrutoquinase. Uma segunda molécula de ATP doa um fosfato de alta energia à frutose-6-fosfato, produzindo frutose-1,6-bisfosfato. Nesta via, a fosfofrutoquinase é uma enzima limitadora da taxa. É ativa quando a concentração de ADP é elevada; é menos ativa quando os níveis de ADP são baixos e aAssim, se houver ATP "suficiente" no sistema, a via torna-se mais lenta. Trata-se de um tipo de inibição do produto final, uma vez que o ATP é o produto final do catabolismo da glicose.
A quarta etapa da glicólise utiliza uma enzima, a aldolase, para clivar o 1,6-bisfosfato em dois isómeros de três carbonos: dihidroxiacetona-fosfato e gliceraldeído-3-fosfato.
Etapa 5 - Na quinta etapa, uma isomerase transforma a dihidroxiacetona-fosfato no seu isómero, o gliceraldeído-3-fosfato. Assim, a via continuará com duas moléculas de um único isómero. Neste ponto da via, há um investimento líquido de energia de duas moléculas de ATP na degradação de uma molécula de glicose.
Segunda metade da glicólise (etapas de libertação de energia)
Até ao momento, a glicólise custou à célula duas moléculas de ATP e produziu duas pequenas moléculas de açúcar de três carbonos. Ambas as moléculas passarão pela segunda metade da via e será extraída energia suficiente para reembolsar as duas moléculas de ATP utilizadas como investimento inicial e produzir um lucro para a célula de duas moléculas de ATP adicionais e duas moléculas de NADH de energia ainda mais elevada.
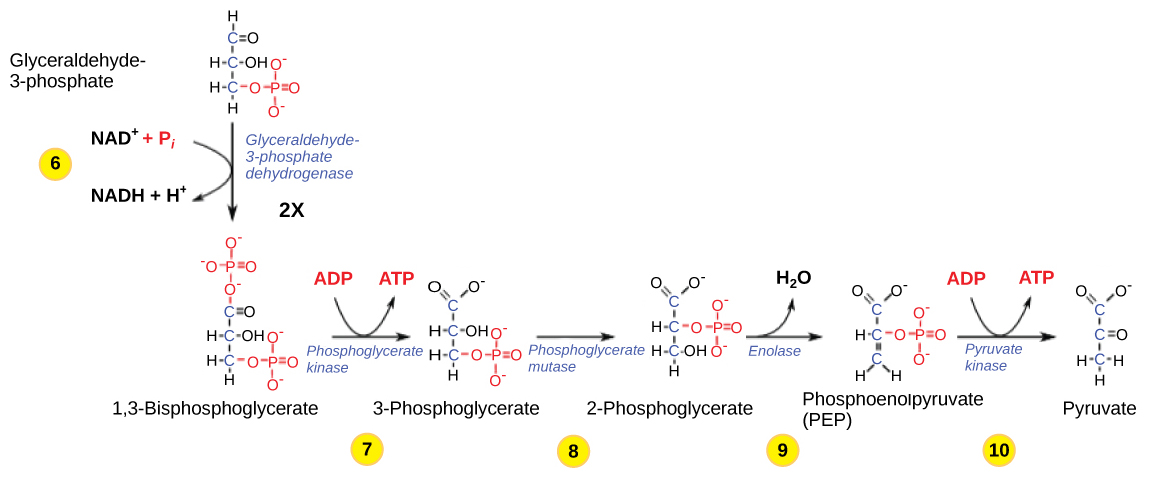
Figura 3: A segunda metade da glicólise envolve a fosforilação sem investimento de ATP (etapa 6) e produz duas moléculas de NADH e quatro moléculas de ATP por glucose.
Etapa 6 - A sexta etapa da glicólise (Figura 3) oxida o açúcar (gliceraldeído-3-fosfato), extraindo electrões de alta energia, que são captados pelo transportador de electrões NAD+, produzindo NADH. O açúcar é então fosforilado pela adição de um segundo grupo fosfato, produzindo 1,3-bisfosfoglicerato.
A continuação da reação depende da disponibilidade da forma oxidada do transportador de electrões, o NAD+. Assim, o NADH tem de ser continuamente oxidado em NAD+ para que esta etapa continue. Se o NAD+ não estiver disponível, a segunda metade da glicólise abranda ou pára. Se houver oxigénio disponível no sistema, o NADH seráNum ambiente sem oxigénio, uma via alternativa (fermentação) pode permitir a oxidação do NADH em NAD+.
Etapa 7: Na sétima etapa, catalisada pela fosfoglicerato quinase (uma enzima cujo nome deriva da reação inversa), o 1,3-bisfosfoglicerato doa um fosfato de alta energia ao ADP, formando uma molécula de ATP (este é um exemplo de fosforilação ao nível do substrato).
Etapa 8: Na oitava etapa, o grupo fosfato remanescente no 3-fosfoglicerato passa do terceiro para o segundo carbono, produzindo 2-fosfoglicerato (um isómero do 3-fosfoglicerato). A enzima que catalisa esta etapa é uma mutase (um tipo de isomerase).
Etapa 9: A enzima enolase catalisa a nona etapa, fazendo com que o 2-fosfoglicerato perca água da sua estrutura; esta é uma reação de desidratação, resultando na formação de uma ligação dupla que aumenta a energia potencial na ligação fosfato restante e produz fosfoenolpiruvato (PEP).
Etapa 10: A última etapa da glicólise é catalisada pela enzima piruvato quinase (neste caso, o nome da enzima deve-se à reação inversa da conversão do piruvato em PEP) e resulta na produção de uma segunda molécula de ATP através da fosforilação ao nível do substrato e do composto ácido pirúvico (ou da sua forma salina, piruvato),uma vez que a enzima pode catalisar tanto reacções de avanço como de retrocesso.
Resultados da glicólise
A glicólise começa com a glicose e termina com duas moléculas de piruvato, um total de quatro moléculas de ATP e duas moléculas de NADH. Duas moléculas de ATP foram utilizadas na primeira metade da via para preparar o anel de seis carbonos para a clivagem, pelo que a célula tem um ganho líquido de duas moléculas de ATP e duas moléculas de NADH para a sua utilização.
Se a célula não conseguir catabolizar mais as moléculas de piruvato, irá obter apenas duas moléculas de ATP a partir de uma molécula de glucose. Os glóbulos vermelhos dos mamíferos maduros não são capazes de respiração aeróbica -Se a glicólise for interrompida, estas células perdem a capacidade de manter as suas bombas de sódio-potássio e acabam por morrer.
O último passo da glicólise não ocorrerá se a piruvato quinase, a enzima que catalisa a formação de piruvato, não estiver disponível em quantidades suficientes. Nesta situação, toda a via glicolítica prosseguirá, mas apenas duas moléculas de ATP serão produzidas na segunda metade. Assim, a piruvato quinase é uma enzima limitadora da taxa de glicólise.
Em resumo: Glicólise
A glicólise é a primeira via utilizada na decomposição da glicose para extrair energia. Foi provavelmente uma das primeiras vias metabólicas a evoluir e é utilizada por quase todos os organismos da Terra. A glicólise consiste em duas partes: a primeira parte prepara o anel de seis carbonos da glicose para ser clivado em dois açúcares de três carbonos.A segunda metade da glicólise extrai ATP e electrões de alta energia dos átomos de hidrogénio e liga-os ao NAD+. Duas moléculas de ATP são investidas na primeira metade e quatro moléculas de ATP são formadas pela fosforilação do substrato durante a segunda metade, o que produz um ganho líquido de duas moléculas de ATP e duas moléculas de NADH para a célula.
A figura 4 mostra todo o processo de glicólise numa só imagem:
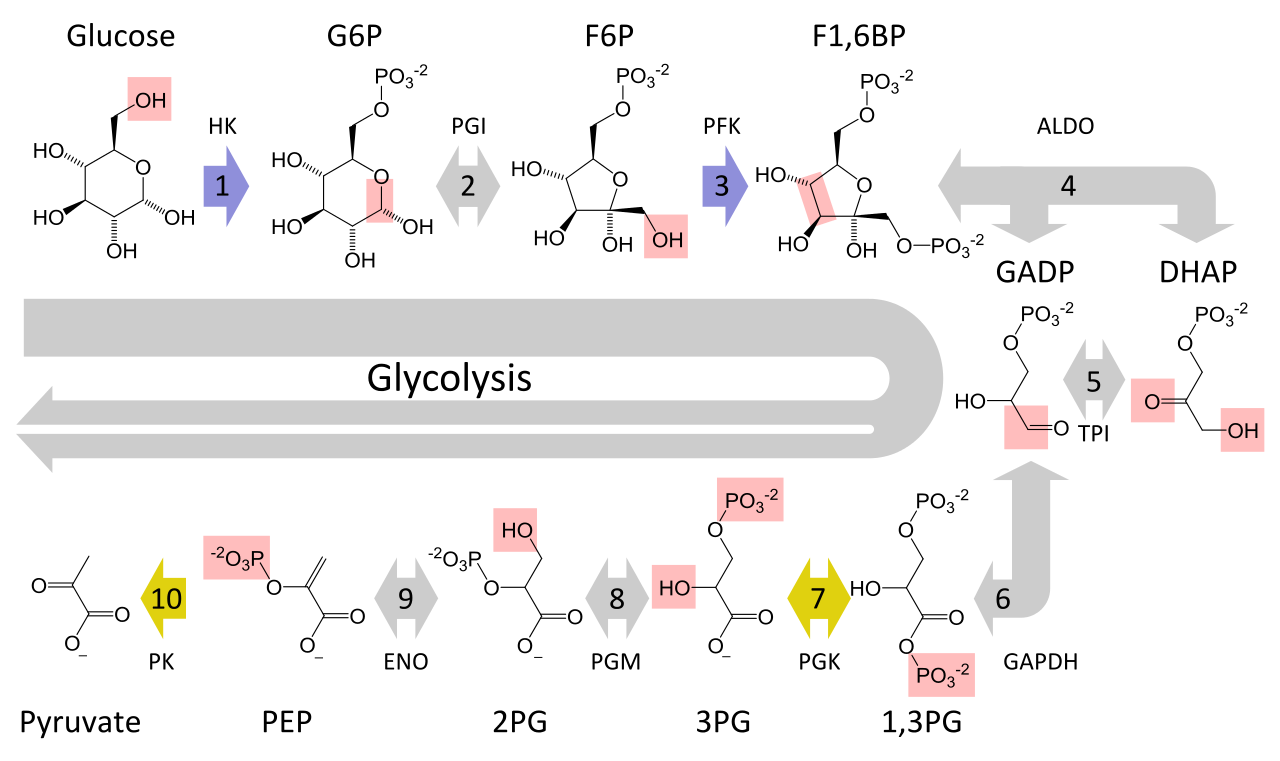
Figura 4: Glicólise
Oxidação do piruvato
Se houver oxigénio disponível, a respiração aeróbica prossegue. Nas células eucarióticas, as moléculas de piruvato produzidas no final da glicólise são transportadas para as mitocôndrias, que são os locais da respiração celular. Aí, o piruvato será transformado num grupo acetilo que será captado e ativado por um composto transportador chamado coenzima A (CoA). O composto resultante é chamado acetil CoA O acetil-CoA é produzido a partir da vitamina B5, o ácido pantoténico. O acetil-CoA pode ser utilizado de várias formas pela célula, mas a sua principal função é fornecer o grupo acetilo derivado do piruvato para a fase seguinte da via do catabolismo da glucose.
Decomposição do piruvato
Para que o piruvato (que é o produto da glicólise) entre no Ciclo do Ácido Cítrico (a via seguinte na respiração celular), tem de sofrer várias alterações. A conversão é um processo em três fases (Figura 5).
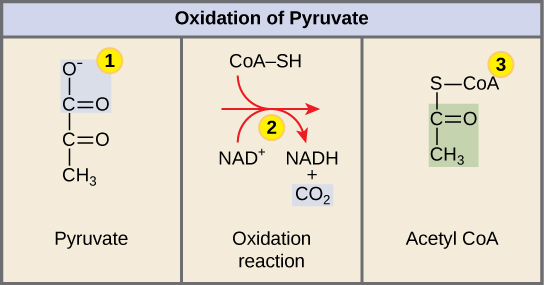
Figura 5. Ao entrar na matriz mitocondrial, um complexo multienzimático converte o piruvato em acetil CoA. No processo, é libertado dióxido de carbono e é formada uma molécula de NADH.
Passo 1: Um grupo carboxilo é removido do piruvato, libertando uma molécula de dióxido de carbono para o meio circundante. O resultado desta etapa é um grupo hidroxietil de dois carbonos ligado à enzima (piruvato desidrogenase). Este é o primeiro dos seis carbonos da molécula de glicose original a ser removido. Esta etapa ocorre duas vezes (lembre-se: há dois moléculas de piruvato produzidas no final da glicólise) por cada molécula de glicose metabolizada; assim, dois dos seis carbonos terão sido removidos no final das duas etapas.
Passo 2: O NAD+ é reduzido a NADH. O grupo hidroxietilo é oxidado a um grupo acetilo e os electrões são captados pelo NAD+, formando NADH. Os electrões de alta energia do NADH serão utilizados mais tarde para gerar ATP.
Passo 3: Um grupo acetilo é transferido para a conenzima A, resultando em acetil CoA. O grupo acetilo ligado à enzima é transferido para a CoA, produzindo uma molécula de acetil CoA.
Note-se que durante a segunda fase do metabolismo da glicose, sempre que um átomo de carbono é removido, liga-se a dois átomos de oxigénio, produzindo dióxido de carbono, um dos principais produtos finais da respiração celular.
Acetil CoA em CO 2
Na presença de oxigénio, o acetil CoA entrega o seu grupo acetil a uma molécula de quatro átomos de carbono, o oxaloacetato, para formar citrato, uma molécula de seis átomos de carbono com três grupos carboxilo; esta via irá recolher o resto da energia extraível do que começou por ser uma molécula de glicose. Ciclo do ácido cítrico .
Em resumo: Oxidação do piruvato
Na presença de oxigénio, o piruvato é transformado num grupo acetilo ligado a uma molécula transportadora de coenzima A. O acetil CoA resultante pode entrar em várias vias, mas, na maioria das vezes, o grupo acetilo é entregue ao ciclo do ácido cítrico para posterior catabolismo. Durante a conversão do piruvato no grupo acetilo, é removida uma molécula de dióxido de carbono e dois electrões de alta energia.O dióxido de carbono representa dois (conversão de duas moléculas de piruvato) dos seis carbonos da molécula de glicose original. Os electrões são captados pelo NAD+, e o NADH transporta os electrões para uma via posterior de produção de ATP. Neste ponto, a molécula de glicose que entrou originalmente na respiração celular foi completamente oxidada. A energia potencial química armazenada na molécula de glicosefoi transferida para transportadores de electrões ou foi utilizada para sintetizar alguns ATP.
Ciclo do ácido cítrico
Tal como a conversão do piruvato em acetil-CoA, o ciclo do ácido cítrico tem lugar na matriz das mitocôndrias. Esta via única é designada por diferentes nomes: ciclo do ácido cítrico (para o primeiro intermediário formado - ácido cítrico ou citrato - quando o acetato se junta ao oxaloacetato), ciclo TCA (uma vez que o ácido cítrico ou citrato e isocitrato são ácidos tricarboxílicos) e ciclo de Krebs , depois de HansKrebs, que identificou pela primeira vez as etapas da via nos anos 30 nos músculos do voo dos pombos.
Quase todas as enzimas do ciclo do ácido cítrico são solúveis, com a única exceção da enzima succinato desidrogenase, que se encontra na membrana interna da mitocôndria. Ao contrário da glicólise, o ciclo do ácido cítrico é um ciclo fechado: a última parte da via regenera o composto utilizado na primeira etapa. As oito etapas do ciclo são uma série de redox, desidratação,reacções de hidratação e descarboxilação que produzem duas moléculas de dióxido de carbono, um GTP/ATP e formas reduzidas de NADH e FADH 2 (Esta é considerada uma via aeróbia porque o NADH e o FADH 2 Se esta transferência não ocorrer, as etapas de oxidação do ciclo do ácido cítrico também não ocorrem. Note-se que o ciclo do ácido cítrico produz diretamente muito pouco ATP e não consome diretamente oxigénio.
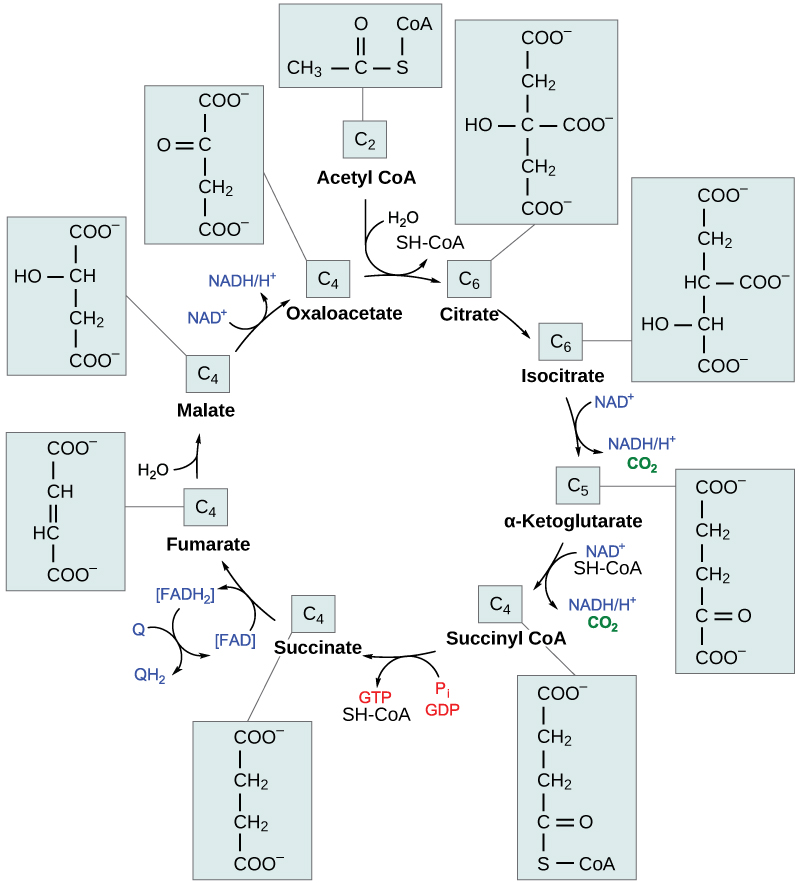
Figura 6. No ciclo do ácido cítrico, o grupo acetil da acetil CoA é ligado a uma molécula de oxaloacetato de quatro carbonos para formar uma molécula de citrato de seis carbonos. Através de uma série de etapas, o citrato é oxidado, libertando duas moléculas de dióxido de carbono por cada grupo acetil introduzido no ciclo. No processo, três moléculas de NAD+ são reduzidas a NADH, uma molécula de FAD é reduzida a FADH 2 Como o produto final do ciclo do ácido cítrico é também o primeiro reagente, o ciclo funciona continuamente na presença de reagentes suficientes. (crédito: modificação do trabalho de "Yikrazuul"/Wikimedia Commons)
Etapas do ciclo do ácido cítrico
Etapa 1: Antes do início da primeira etapa, deve ocorrer a oxidação do piruvato. Em seguida, inicia-se a primeira etapa do ciclo: trata-se de uma etapa de condensação, que combina o grupo acetilo de dois carbonos com uma molécula de oxaloacetato de quatro carbonos para formar uma molécula de seis carbonos de citrato A CoA liga-se a um grupo sulfidrilo (-SH) e difunde-se para eventualmente se combinar com outro grupo acetilo. Esta etapa é irreversível porque é altamente exergónica. A velocidade desta reação é controlada por feedback negativo e pela quantidade de ATP disponível. Se os níveis de ATP aumentarem, a velocidade desta reação diminui. Se o ATP for escasso, a velocidade aumenta.
Etapa 2: Na segunda etapa, o citrato perde uma molécula de água e ganha outra, uma vez que o citrato é convertido no seu isómero, isocitrato .
Etapa 3: Na terceira etapa, o isocitrato é oxidado, produzindo uma molécula de cinco carbonos, α-cetoglutarato juntamente com uma molécula de CO 2 e dois electrões, que reduzem o NAD+ a NADH. Esta etapa é também regulada por um feedback negativo do ATP e do NADH, e por um efeito positivo do ADP.
Etapas 3 e 4: As etapas três e quatro são etapas de oxidação e descarboxilação, que libertam electrões que reduzem o NAD+ a NADH e libertam grupos carboxilo que formam CO 2 O α-cetoglutarato é o produto da terceira etapa, e um succinilo O CoA liga-se ao grupo succinil para formar succinil CoA. A enzima que catalisa a etapa quatro é regulada pela inibição por retroação do ATP, succinil CoA e NADH.
Etapa 5: Na quinta etapa, um grupo fosfato é substituído pela coenzima A e forma-se uma ligação de alta energia, que é utilizada na fosforilação ao nível do substrato (durante a conversão do grupo succinilo em succinato) para formar guanina trifosfato (GTP) ou ATP. Existem duas formas da enzima, denominadas isoenzimas, para esta etapa, consoante o tipo de tecido animal em que se encontramUma forma encontra-se nos tecidos que utilizam grandes quantidades de ATP, como o coração e o músculo esquelético. Esta forma produz ATP. A segunda forma da enzima encontra-se nos tecidos que têm um elevado número de vias anabólicas, como o fígado. Esta forma produz GTP. O GTP é energeticamente equivalente ao ATP; no entanto, a sua utilização é mais restrita. Em particular, a síntese proteica utiliza principalmente GTP.
Etapa 6 - A etapa 6 é um processo de desidratação que converte succinato em fumarato Dois átomos de hidrogénio são transferidos para o FAD, produzindo FADH 2 A energia contida nos electrões destes átomos é insuficiente para reduzir o NAD+ mas suficiente para reduzir o FAD. Ao contrário do NADH, este transportador permanece ligado à enzima e transfere os electrões diretamente para a cadeia de transporte de electrões. Este processo é possível graças à localização da enzima que catalisa esta etapa no interior da membrana interna da mitocôndria.
Etapa 7: durante a etapa 7, adiciona-se água ao fumarato e malato A última etapa do ciclo do ácido cítrico regenera o oxaloacetato através da oxidação do malato, sendo produzida outra molécula de NADH.
Pode clicar em cada passo do ciclo do ácido cítrico aqui.Produtos do ciclo do ácido cítrico
Dois átomos de carbono entram no ciclo do ácido cítrico a partir de cada grupo acetil, representando quatro dos seis carbonos de uma molécula de glicose. Duas moléculas de dióxido de carbono são libertadas em cada volta do ciclo; no entanto, estas não contêm necessariamente os átomos de carbono adicionados mais recentemente. Os dois átomos de carbono acetil serão eventualmente libertados em voltas posteriores do ciclo; assim, todos os seis átomos de carbonoCada volta do ciclo forma três moléculas de NADH e uma de FADH 2 Estes transportadores ligam-se à última porção da respiração aeróbica para produzir moléculas de ATP. Em cada ciclo é também produzido um GTP ou ATP. Vários dos compostos intermédios do ciclo do ácido cítrico podem ser utilizados na síntese de aminoácidos não essenciais; por conseguinte, o ciclo é anfibólico (tanto catabólico como anabólico).
Em resumo: Ciclo do ácido cítrico
O ciclo do ácido cítrico é uma série de reacções redox e de descarboxilação que removem electrões de alta energia e dióxido de carbono. Os electrões são temporariamente armazenados em moléculas de NADH e FADH 2 Uma molécula de GTP ou ATP é produzida por fosforilação ao nível do substrato em cada volta do ciclo. Não há comparação da via cíclica com uma via linear.
Cadeia de transporte de electrões
Acabou de ler sobre duas vias da respiração celular - a glicólise e o ciclo do ácido cítrico - que geram ATP. No entanto, a maior parte do ATP gerado durante o catabolismo aeróbio da glicose não é gerado diretamente por estas vias, mas sim por um processo que começa com a movimentação de electrões através de uma série de transportadores de electrões que sofrem reacções redox: a cadeia de transporte de electrões Isto faz com que os iões de hidrogénio se acumulem no espaço da matriz, formando-se assim um gradiente de concentração no qual os iões de hidrogénio se difundem para fora do espaço da matriz, passando pela ATP sintase. A corrente de iões de hidrogénio alimenta a ação catalítica da ATP sintase, que fosforila o ADP, produzindo ATP.
Cadeia de transporte de electrões
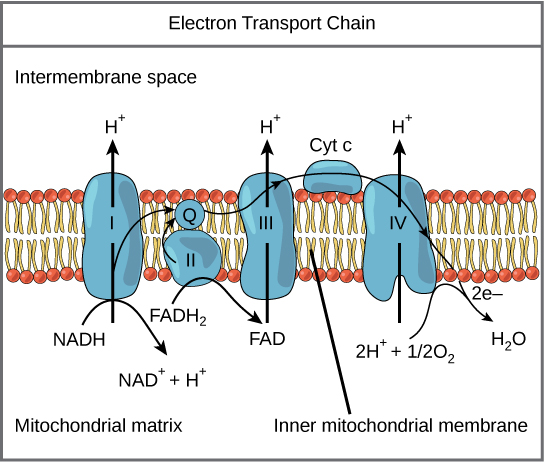
Figura 7: A cadeia de transporte de electrões é uma série de transportadores de electrões integrados na membrana mitocondrial interna que transporta os electrões do NADH e do FADH 2 No processo, os protões são bombeados da matriz mitocondrial para o espaço intermembranar e o oxigénio é reduzido para formar água.
A cadeia de transporte de electrões (Figura 7) é o último componente da respiração aeróbia e é a única parte do metabolismo da glicose que utiliza o oxigénio atmosférico. O oxigénio difunde-se continuamente nas plantas; nos animais, entra no corpo através do sistema respiratório. O transporte de electrões é uma série de reacções redox que se assemelham a uma corrida de estafetas ou a uma brigada de baldes, na medida em que os electrões passam rapidamente deExistem quatro complexos compostos por proteínas, assinalados de I a IV na Figura 7, e a agregação destes quatro complexos, juntamente com os transportadores de electrões acessórios móveis associados, é designada por cadeia de transporte de electrões. A cadeia de transporte de electrões está presente em múltiplosA cadeia de transporte de electrões dos procariotas pode não necessitar de oxigénio, uma vez que alguns vivem em condições anaeróbias. A caraterística comum a todas as cadeias de transporte de electrões é a presença de uma bomba de protões para criar um gradiente de protões através de uma membrana.
Complexo I
Para começar, dois electrões são transportados para o primeiro complexo a bordo do NADH. Este complexo, denominado I, é composto pelo mononucleótido de flavina (FMN) e por uma proteína que contém ferro-enxofre (Fe-S). O FMN, que é derivado da vitamina B 2 A riboflavina, também designada por riboflavina, é um dos vários grupos prostéticos ou co-factores da cadeia de transporte de electrões. grupo protético Os grupos prostéticos são moléculas orgânicas ou inorgânicas não peptídicas ligadas a uma proteína que facilitam a sua função; os grupos prostéticos incluem as co-enzimas, que são os grupos prostéticos das enzimas. A enzima do complexo I é a NADH desidrogenase e é uma proteína muito grande, contendo 45 cadeias de aminoácidos. O complexo I pode bombear quatroiões de hidrogénio através da membrana, da matriz para o espaço intermembranar, e é desta forma que o gradiente de iões de hidrogénio é estabelecido e mantido entre os dois compartimentos separados pela membrana mitocondrial interna.
Q e Complexo II
O complexo II recebe diretamente FADH 2 que não passa pelo complexo I. O composto que liga o primeiro e o segundo complexos ao terceiro é ubiquinona (Q) A molécula Q é lipossolúvel e move-se livremente através do núcleo hidrofóbico da membrana. Uma vez reduzida, (QH 2 ), a ubiquinona entrega os seus electrões ao complexo seguinte da cadeia de transporte de electrões. Q recebe os electrões provenientes do NADH do complexo I e os electrões provenientes do FADH 2 do complexo II, incluindo a succinato desidrogenase. Esta enzima e a FADH 2 Uma vez que estes electrões não energizam a bomba de protões no primeiro complexo, são produzidas menos moléculas de ATP a partir do FADH 2 O número de moléculas de ATP finalmente obtidas é diretamente proporcional ao número de protões bombeados através da membrana mitocondrial interna.
Complexo III
O terceiro complexo é composto pelo citocromo b, por outra proteína Fe-S, pelo centro de Rieske (centro 2Fe-2S) e pelas proteínas do citocromo c; este complexo é também designado por citocromo oxidoredutase. As proteínas do citocromo têm um grupo prostético de heme. A molécula de heme é semelhante à molécula de heme da hemoglobina, mas transporta electrões e não oxigénio. Consequentemente, o ião de ferro no seu núcleo é reduzido e oxidado à medida queAs moléculas de heme nos citocromos têm características ligeiramente diferentes devido aos efeitos das diferentes proteínas que as ligam, dando características ligeiramente diferentes a cada complexo. O complexo III bombeia protões através da membrana e passa os seus electrões para o citocromo c paratransporte para o quarto complexo de proteínas e enzimas (o citocromo c é o aceitador de electrões de Q; no entanto, enquanto Q transporta pares de electrões, o citocromo c só pode aceitar um de cada vez).
Complexo IV
O quarto complexo é composto pelas proteínas do citocromo c, a e a 3 Este complexo contém dois grupos heme (um em cada um dos dois citocromos, a, e a 3 ) e três iões de cobre (um par de Cu A e um Cu B no citocromo a 3 Os citocromos mantêm uma molécula de oxigénio muito apertada entre os iões de ferro e de cobre até que o oxigénio seja completamente reduzido. O oxigénio reduzido apanha então dois iões de hidrogénio do meio circundante para formar água (H 2 O) A remoção dos iões de hidrogénio do sistema contribui para o gradiente iónico utilizado no processo de quimiosmose.
Quimiosmose
Na quimiosmose, a energia livre da série de reacções redox que acabámos de descrever é utilizada para bombear iões de hidrogénio (protões) através da membrana. A distribuição desigual dos iões H+ através da membrana estabelece gradientes de concentração e eléctricos (ou seja, um gradiente eletroquímico), devido à carga positiva dos iões de hidrogénio e à sua agregação num dos lados da membrana.
Se a membrana estivesse aberta à difusão dos iões de hidrogénio, os iões tenderiam a difundir-se de volta para a matriz, impulsionados pelo seu gradiente eletroquímico. Recorde-se que muitos iões não podem difundir-se através das regiões não polares das membranas fosfolipídicas sem a ajuda de canais iónicos. Do mesmo modo, os iões de hidrogénio no espaço da matriz só podem passar através da membrana mitocondrial interna através de umEsta proteína complexa actua como um minúsculo gerador, que gira pela força dos iões de hidrogénio que se difundem através dela, descendo o seu gradiente eletroquímico. A rotação das peças desta máquina molecular facilita a adição de um fosfato ao ADP, formando ATP, utilizando a energia potencial do gradiente de iões de hidrogénio.