- O modelo de contração do filamento deslizante
- ATP e Contração Muscular
- Fontes de ATP
- Relaxamento de um músculo esquelético
- Força muscular
- Perguntas de auto-verificação
Objectivos de aprendizagem
- Descrever os componentes envolvidos numa contração muscular
- Explicar como os músculos se contraem e relaxam
- Descrever o modelo de filamentos deslizantes da contração muscular
A sequência de eventos que resultam na contração de uma fibra muscular individual começa com um sinal - o neurotransmissor ACh - do neurónio motor que inerva essa fibra. A membrana local da fibra despolariza-se com a entrada de iões de sódio (Na+) carregados positivamente, desencadeando um potencial de ação que se propaga para o resto da membrana que se despolariza, incluindo os túbulos T. Isto desencadeiaA libertação de iões de cálcio (Ca++) a partir do armazenamento no retículo sarcoplasmático (SR). O Ca++ inicia então a contração, que é sustentada pelo ATP (Figura 1). Enquanto os iões Ca++ permanecerem no sarcoplasma para se ligarem à troponina, o que mantém os locais de ligação à actina "desprotegidos", e enquanto o ATP estiver disponível para conduzir o ciclo da ponte cruzada e a tração dos filamentos de actina pela miosina, o músculoA fibra continuará a encurtar até um limite anatómico.
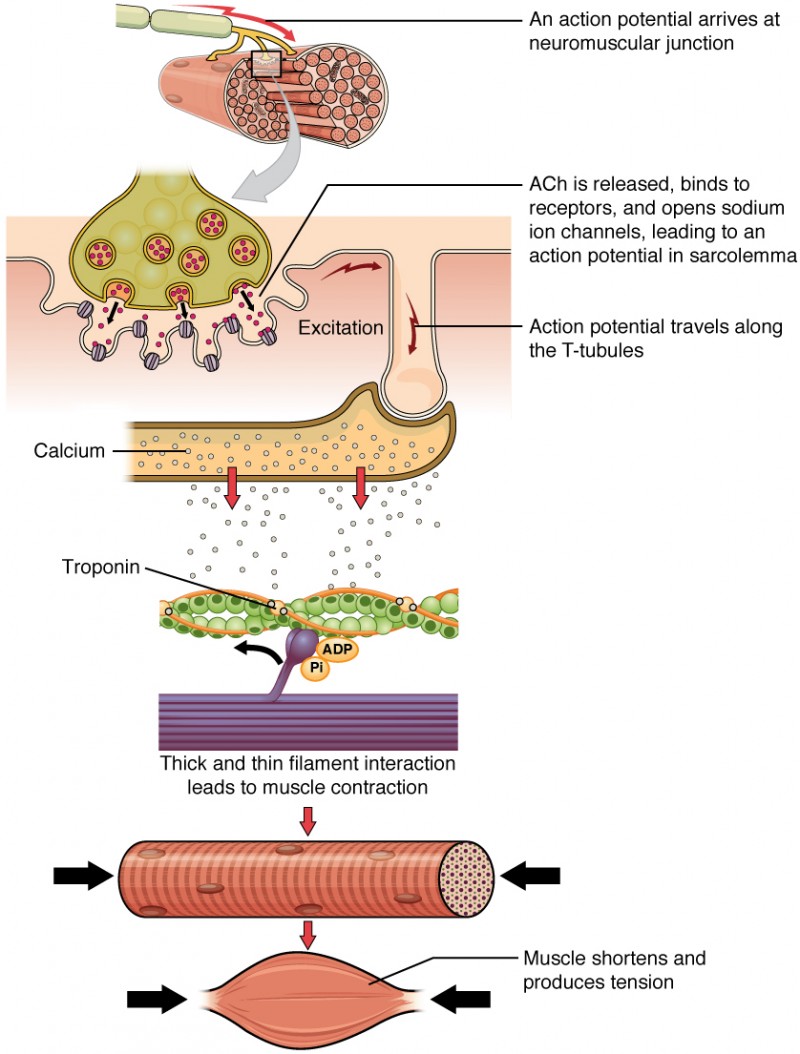
Figura 1: Contração de uma fibra muscular Enquanto os iões Ca++ permanecerem no sarcoplasma para se ligarem à troponina, e enquanto houver ATP disponível, a fibra muscular continuará a encurtar.
A contração muscular pára normalmente quando a sinalização do neurónio motor termina, o que repolariza o sarcolema e os túbulos em T e fecha os canais de cálcio dependentes da voltagem no RE. Os iões Ca++ são então bombeados de volta para o RE, o que faz com que a tropomiosina volte a proteger (ou volte a cobrir) os locais de ligação nos filamentos de actina. Um músculo também pode parar de se contrair quando fica sem ATP e se cansa(Figura 2).
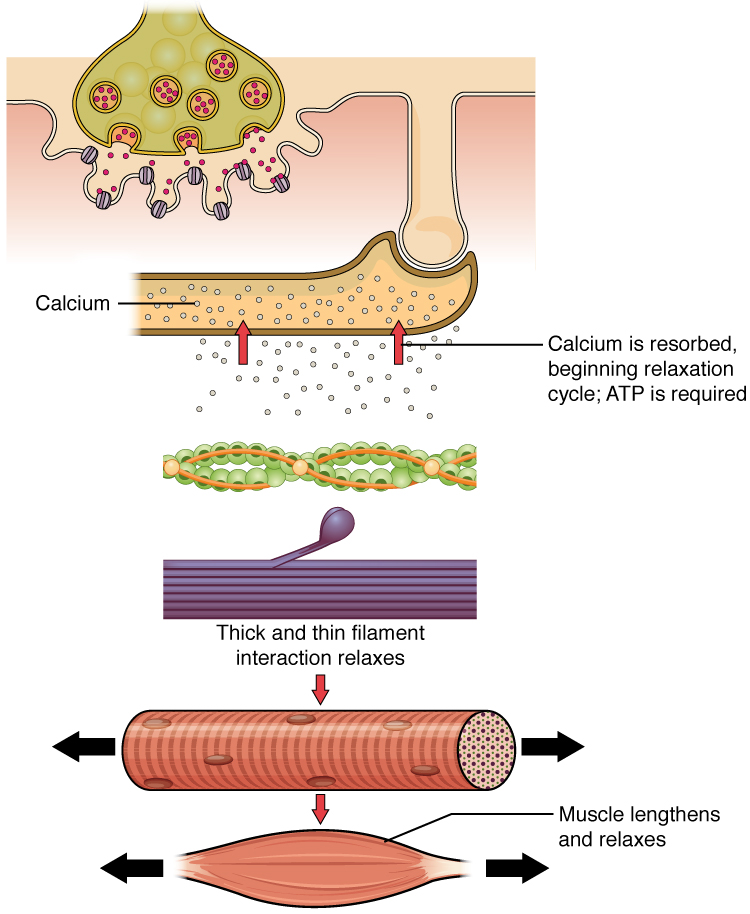
Figura 2. Relaxamento de uma fibra muscular. Os iões Ca++ são bombeados de volta para o RE, o que faz com que a tropomiosina volte a proteger os locais de ligação nos filamentos de actina. Um músculo também pode parar de se contrair quando fica sem ATP e entra em fadiga.
A libertação de iões de cálcio inicia as contracções musculares. Assista a este vídeo para saber mais sobre o papel do cálcio. a) O que são os "túbulos em T" e qual é o seu papel? b) Descreva como é que os locais de ligação da actina são disponibilizados para a formação de pontes cruzadas com as cabeças de miosina durante a contração.Os eventos moleculares do encurtamento da fibra muscular ocorrem nos sarcómeros da fibra (ver Figura 3). A contração de uma fibra muscular estriada ocorre quando os sarcómeros, dispostos linearmente nas miofibrilas, encurtam à medida que as cabeças de miosina puxam os filamentos de actina.
A região onde os filamentos finos e grossos se sobrepõem tem uma aparência densa, pois há pouco espaço entre os filamentos. Esta zona onde os filamentos finos e grossos se sobrepõem é muito importante para a contração muscular, pois é o local onde se inicia o movimento dos filamentos. Os filamentos finos, ancorados nas suas extremidades pelos discos Z, não se estendem completamente para a região central que contém apenas filamentos grossosUma miofibrila é composta por muitos sarcómeros ao longo do seu comprimento; assim, as miofibrilas e as células musculares contraem-se à medida que os sarcómeros se contraem.
O modelo de contração do filamento deslizante
Quando sinalizada por um neurónio motor, uma fibra muscular esquelética contrai-se à medida que os filamentos finos são puxados e depois deslizam pelos filamentos espessos dentro dos sarcómeros da fibra. Este processo é conhecido como o modelo de filamentos deslizantes da contração muscular (Figura 3). O deslizamento só pode ocorrer quando os locais de ligação da miosina nos filamentos de actina são expostos por uma série de passos que começam com a entrada de Ca++ noo sarcoplasma.
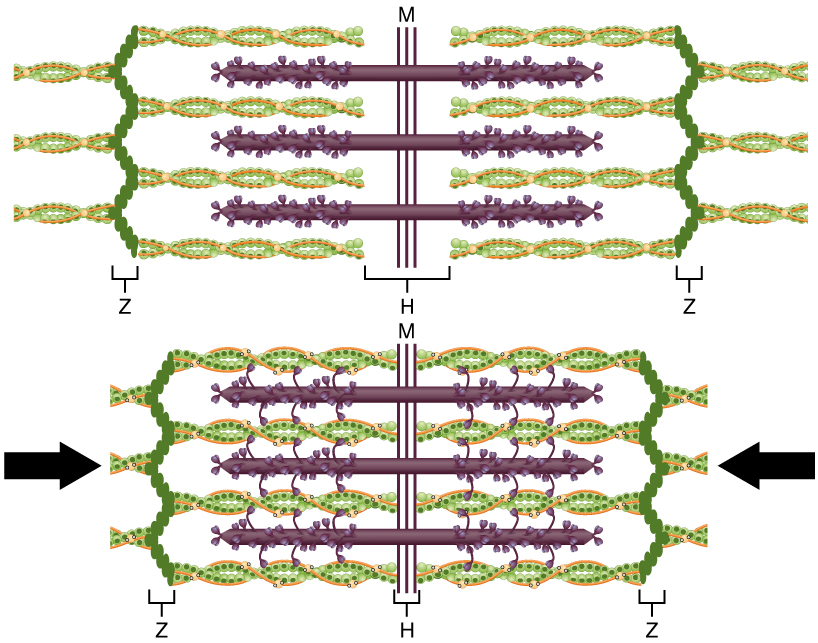
Figura 3: O modelo de filamento deslizante da contração muscular. Quando um sarcómero se contrai, as linhas Z aproximam-se umas das outras e a banda I torna-se mais pequena. A banda A mantém a mesma largura. Na contração total, os filamentos finos e grossos sobrepõem-se.
A tropomiosina é uma proteína que se enrola à volta das cadeias do filamento de actina e cobre os locais de ligação da miosina para impedir que a actina se ligue à miosina. A tropomiosina liga-se à troponina para formar um complexo troponina-tropomiosina. O complexo troponina-tropomiosina impede que as "cabeças" da miosina se liguem aos locais activos nos microfilamentos de actina. A troponina também tem um local de ligação para iões Ca++.
Para iniciar a contração muscular, a tropomiosina tem de expor o local de ligação da miosina num filamento de actina para permitir a formação de pontes cruzadas entre os microfilamentos de actina e de miosina. O primeiro passo no processo de contração é a ligação do Ca++ à troponina, para que a tropomiosina possa deslizar para longe dos locais de ligação nos filamentos de actina, o que permite que as cabeças de miosina se liguem a estes locais de ligação expostosOs filamentos finos são então puxados pelas cabeças de miosina para deslizarem para além dos filamentos grossos em direção ao centro do sarcómero. Mas cada cabeça só pode puxar uma distância muito curta antes de atingir o seu limite e tem de ser "rearmada" antes de poder puxar novamente, um passo que requer ATP.
ATP e Contração Muscular
Para que os filamentos finos continuem a deslizar sobre os filamentos espessos durante a contração muscular, as cabeças de miosina têm de puxar a actina nos locais de ligação, desprender-se, voltar a prender-se, ligar-se a mais locais de ligação, puxar, desprender-se, voltar a prender-se, etc. Este movimento repetido é conhecido como o ciclo das pontes cruzadas. Este movimento das cabeças de miosina é semelhante ao dos remos quando um indivíduo rema um barco: a pá dos remos (a miosinaCada ciclo requer energia, e a ação das cabeças de miosina nos sarcómeros que puxam repetidamente os filamentos finos também requer energia, que é fornecida pelo ATP.
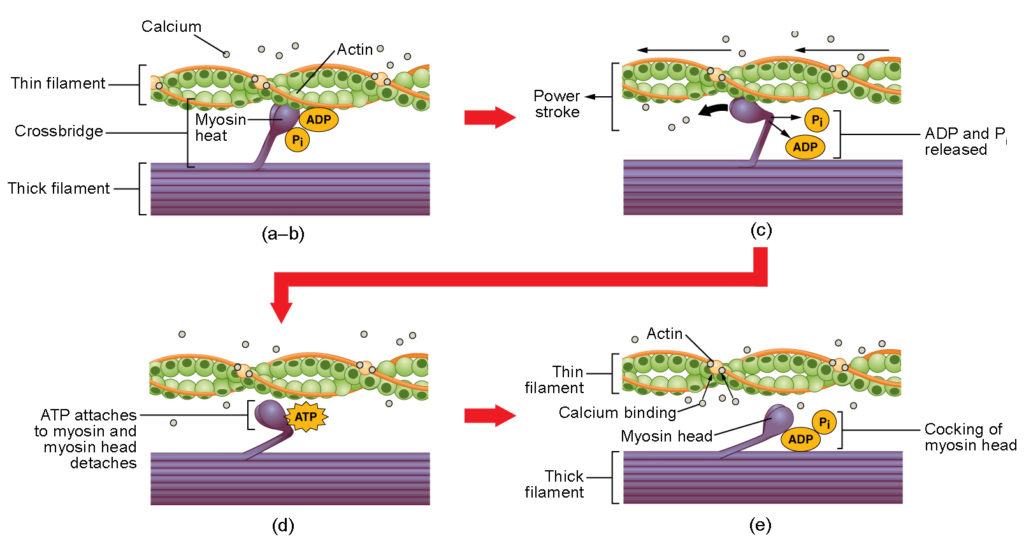
Figura 4. Contração do músculo esquelético. (a) O sítio ativo da actina é exposto à medida que o cálcio se liga à troponina. (b) A cabeça da miosina é atraída pela actina e a miosina liga-se à actina no seu sítio de ligação à actina, formando a ponte cruzada. (c) Durante o ciclo de força, o fosfato gerado no ciclo de contração anterior é libertado, o que faz com que a cabeça da miosina gire em direção ao centro do sarcómero, após o que o ADP e o(d) Uma nova molécula de ATP liga-se à cabeça da miosina, fazendo com que a ponte cruzada se desprenda. (e) A cabeça da miosina hidrolisa o ATP em ADP e fosfato, o que faz com que a miosina volte à posição de cócoras.
A formação de pontes cruzadas ocorre quando a cabeça da miosina se liga à actina enquanto o difosfato de adenosina (ADP) e o fosfato inorgânico (P i ) ainda estão ligados à miosina (Figura 4a,b). i é então libertada, fazendo com que a miosina forme uma ligação mais forte à actina, após o que a cabeça da miosina se move em direção à linha M, puxando a actina consigo. À medida que a actina é puxada, os filamentos movem-se cerca de 10 nm em direção à linha M. Este movimento é designado por curso de potência Na ausência de ATP, a cabeça da miosina não se desprende da actina.
Uma parte da cabeça da miosina liga-se ao local de ligação na actina, mas a cabeça tem outro local de ligação para o ATP. A ligação do ATP faz com que a cabeça da miosina se separe da actina (Figura 4d). Depois de isto acontecer, o ATP é convertido em ADP e P i pelo intrínseco ATPase A energia libertada durante a hidrólise do ATP altera o ângulo da cabeça da miosina para uma posição inclinada (Figura 4e). A cabeça da miosina está agora em posição para continuar o movimento.
Quando a cabeça da miosina está inclinada, a miosina está numa configuração de alta energia. Esta energia é gasta à medida que a cabeça da miosina se move através do curso de força e, no final do curso de força, a cabeça da miosina está numa posição de baixa energia. Após o curso de força, o ADP é libertado; no entanto, a ponte cruzada formada ainda está no lugar e a actina e a miosina estão ligadas. Enquanto o ATP estiver disponível, eleprontamente se liga à miosina, o ciclo da ponte cruzada pode recomeçar e a contração muscular pode continuar.
Repare que cada filamento espesso de cerca de 300 moléculas de miosina tem várias cabeças de miosina e que muitas pontes cruzadas se formam e quebram continuamente durante a contração muscular. Multiplique isto por todos os sarcómeros de uma miofibrila, por todas as miofibrilas de uma fibra muscular e por todas as fibras musculares de um músculo esquelético e poderá compreender por que razão é necessária tanta energia (ATP) para manter os músculos esqueléticosNa verdade, é a perda de ATP que resulta no rigor mortis observado logo após a morte de uma pessoa. Não sendo possível produzir mais ATP, não há ATP disponível para as cabeças de miosina se separarem dos locais de ligação à actina, pelo que as pontes cruzadas permanecem no lugar, causando a rigidez nos músculos esqueléticos.
Fontes de ATP
O ATP fornece a energia necessária para a contração muscular. Para além do seu papel direto no ciclo das pontes cruzadas, o ATP também fornece a energia para as bombas de transporte ativo de Ca++ no RE. A contração muscular não ocorre sem quantidades suficientes de ATP. A quantidade de ATP armazenada no músculo é muito baixa, apenas suficiente para alimentar alguns segundos de contracções. À medida que é decomposto, o ATPExistem três mecanismos pelos quais o ATP pode ser regenerado: o metabolismo da creatina fosfato, a glicólise anaeróbica, a fermentação e a respiração aeróbica.
Fosfato de creatina É uma molécula que pode armazenar energia nas suas ligações de fosfato. Num músculo em repouso, o excesso de ATP transfere a sua energia para a creatina, produzindo ADP e fosfato de creatina. Esta actua como uma reserva de energia que pode ser utilizada para criar rapidamente mais ATP. Quando o músculo começa a contrair-se e necessita de energia, o fosfato de creatina transfere o seu fosfato de volta para o ADP para formar ATP e creatina. Esta reação écatalisada pela enzima creatina quinase e ocorre muito rapidamente; assim, o ATP derivado do fosfato de creatina alimenta os primeiros segundos da contração muscular. No entanto, o fosfato de creatina só pode fornecer aproximadamente 15 segundos de energia, altura em que tem de ser utilizada outra fonte de energia (Figura 5).
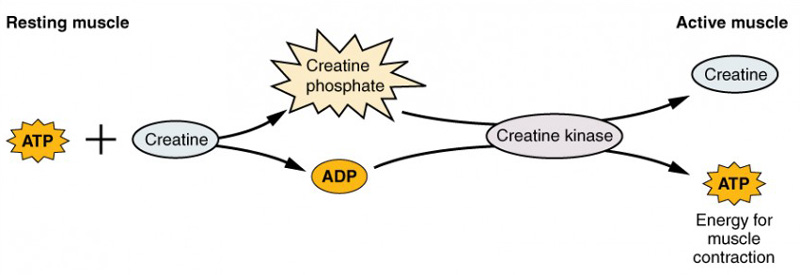
Figura 5: Metabolismo muscular. No músculo em repouso é armazenado algum ATP que, quando se inicia a contração, é consumido em segundos. É gerado mais ATP a partir do fosfato de creatina durante cerca de 15 segundos.
À medida que o ATP produzido pelo fosfato de creatina se esgota, os músculos recorrem à glicólise como fonte de ATP. Glicólise é um processo anaeróbico (não dependente de oxigénio) que decompõe a glicose (açúcar) para produzir ATP; no entanto, a glicólise não pode gerar ATP tão rapidamente como a creatina fosfato. Assim, a mudança para a glicólise resulta numa taxa mais lenta de disponibilidade de ATP para o músculo. O açúcar utilizado na glicólise pode ser fornecido pela glicose sanguínea ou pelo metabolismo do glicogénio armazenado no músculo. A decomposição deuma molécula de glucose produz dois ATP e duas moléculas de ácido pirúvico que pode ser utilizado na respiração aeróbica ou, quando os níveis de oxigénio são baixos, convertido em ácido lático (Figura 6).
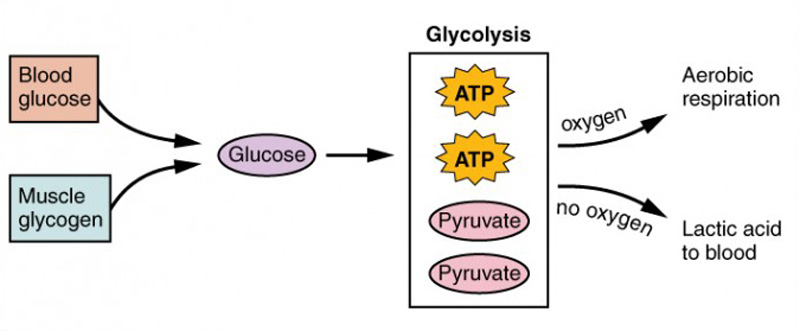
Figura 6: Glicólise e respiração aeróbica. Cada molécula de glicose produz dois ATP e duas moléculas de ácido pirúvico, que pode ser utilizado na respiração aeróbica ou convertido em ácido lático. Se não houver oxigénio disponível, o ácido pirúvico é convertido em ácido lático, o que pode contribuir para a fadiga muscular. Isto ocorre durante o exercício extenuante, quando são necessárias grandes quantidades de energia, mas o oxigénio não pode ser suficientemente fornecido ao músculo.
Se houver oxigénio disponível, o ácido pirúvico é utilizado na respiração aeróbia. No entanto, se não houver oxigénio disponível, o ácido pirúvico é convertido em ácido lático Esta conversão permite a reciclagem da enzima NAD+ a partir do NADH, que é necessária para que a glicólise continue. Isto ocorre durante o exercício extenuante, quando são necessárias grandes quantidades de energia, mas o oxigénio não pode ser suficientemente fornecido ao músculo. A glicólise em si não pode ser sustentada durante muito tempo (aproximadamente 1 minuto de atividade muscular), mas é útilIsto deve-se ao facto de a glicólise não utilizar a glicose de forma muito eficiente, produzindo um ganho líquido de dois ATP por molécula de glicose e o produto final de ácido lático, que pode contribuir para a fadiga muscular à medida que se acumula.
Respiração aeróbica é a decomposição da glucose ou de outros nutrientes na presença de oxigénio (O 2 Aproximadamente 95 por cento do ATP necessário para os músculos em repouso ou moderadamente activos é fornecido pela respiração aeróbica, que tem lugar nas mitocôndrias. Os factores de produção da respiração aeróbica incluem a glicose que circula na corrente sanguínea, o ácido pirúvico e os ácidos gordos. A respiração aeróbica é muito mais eficiente do que a glicólise anaeróbica, produzindoNo entanto, a respiração aeróbica não pode ser sustentada sem um fornecimento constante de O 2 Para compensar, os músculos armazenam uma pequena quantidade de oxigénio em excesso em proteínas chamadas mioglobina, o que permite contracções musculares mais eficientes e menos fadiga. O treino aeróbico também aumenta a eficiência do sistema circulatório, de modo a que o O 2 pode ser fornecido aos músculos durante períodos de tempo mais longos.
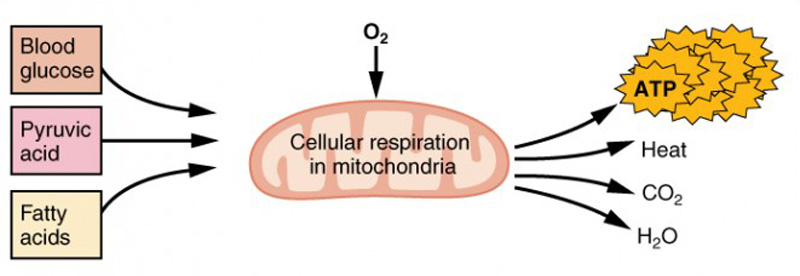
Figura 7: Respiração celular. A respiração aeróbica é a decomposição da glicose na presença de oxigénio (O 2 Cerca de 95% do ATP necessário para os músculos em repouso ou moderadamente activos é fornecido pela respiração aeróbica, que tem lugar nas mitocôndrias.
A fadiga muscular ocorre quando um músculo já não se consegue contrair em resposta a sinais do sistema nervoso. As causas exactas da fadiga muscular não são totalmente conhecidas, embora certos factores tenham sido correlacionados com a diminuição da contração muscular que ocorre durante a fadiga. O ATP é necessário para a contração muscular normal e, à medida que as reservas de ATP são reduzidas, a função muscular pode diminuir. Isto pode ser maisA acumulação de ácido lático pode reduzir o pH intracelular, afectando a atividade enzimática e proteica. Os desequilíbrios nos níveis de Na+ e K+ resultantes da despolarização da membrana podem perturbar o fluxo de Ca++ para fora do RE. Os longos períodos de exercício sustentado podem danificar o RE e o sarcolema, resultando numa regulação deficiente do Ca++.
A atividade muscular intensa resulta numa dívida de oxigénio O oxigénio é necessário para repor os níveis de ATP e de fosfato de creatina, para converter o ácido lático em ácido pirúvico e, no fígado, para converter o ácido lático em glicose ou glicogénio. Outros sistemas utilizados durante o exercício também necessitam de oxigénio, e todos estes processos combinados resultam no aumento daAté que o débito de oxigénio seja satisfeito, o consumo de oxigénio é elevado, mesmo após a interrupção do exercício.
Relaxamento de um músculo esquelético
O relaxamento das fibras musculares esqueléticas e, em última análise, do músculo esquelético, começa com o neurónio motor, que deixa de libertar o seu sinal químico, a ACh, na sinapse no NMJ. A fibra muscular repolariza-se, o que fecha os portões no SR onde o Ca++ estava a ser libertado. As bombas accionadas por ATP deslocam o Ca++ do sarcoplasma para o SR. Isto resulta na "blindagem" da actina-Sem a capacidade de formar pontes cruzadas entre os filamentos finos e grossos, a fibra muscular perde a tensão e relaxa.
Força muscular
O número de fibras musculares esqueléticas num determinado músculo é determinado geneticamente e não se altera. A força muscular está diretamente relacionada com a quantidade de miofibrilas e sarcómeros existentes em cada fibra. Factores como as hormonas e o stress (e os esteróides anabolizantes artificiais), que actuam no músculo, podem aumentar a produção de sarcómeros e miofibrilas dentro das fibras musculares, uma alteração denominadaA hipertrofia, que resulta no aumento da massa e do volume de um músculo esquelético, e a diminuição do uso de um músculo esquelético resultam em atrofia, em que o número de sarcómeros e de miofibrilas desaparece (mas não o número de fibras musculares). É comum que um membro engessado apresente músculos atrofiados quando o gesso é retirado, e certas doenças, como a poliomielite, apresentam músculos atrofiados.
Perturbações do sistema muscular
A distrofia muscular de Duchenne (DMD) é um enfraquecimento progressivo dos músculos esqueléticos. É uma das várias doenças coletivamente designadas por "distrofia muscular". A DMD é causada pela falta da proteína distrofina, que ajuda os filamentos finos das miofibrilas a ligarem-se ao sarcolema. Sem distrofina suficiente, as contracções musculares provocam a rutura do sarcolema, causando um influxo de Ca++,Com o tempo, à medida que os danos musculares se acumulam, perde-se massa muscular e desenvolvem-se maiores incapacidades funcionais.
A DMD é uma doença hereditária causada por uma anomalia no cromossoma X. Afecta principalmente os homens e é normalmente diagnosticada na primeira infância. A DMD surge normalmente como uma dificuldade de equilíbrio e de movimento, progredindo depois para uma incapacidade de andar. Continua a progredir no corpo, das extremidades inferiores para a parte superior do corpo, onde afecta os músculos responsáveis pela respiração eA circulação sanguínea, que acaba por provocar a morte por insuficiência respiratória, não ultrapassa normalmente os 20 anos de idade.
Uma vez que a DMD é causada por uma mutação no gene que codifica a distrofina, pensou-se que a introdução de mioblastos saudáveis nos doentes poderia ser um tratamento eficaz. Os mioblastos são as células embrionárias responsáveis pelo desenvolvimento muscular e, idealmente, seriam portadores de genes saudáveis que poderiam produzir a distrofina necessária para uma contração muscular normal.Uma abordagem recente envolveu a tentativa de aumentar a produção muscular de utrofina, uma proteína semelhante à distrofina que pode ser capaz de assumir o papel da distrofina e impedir a ocorrência de danos celulares.
Perguntas de auto-verificação
Faça o teste abaixo para verificar a sua compreensão da Contração e Relaxamento das Fibras Musculares: